In the dynamic world of veterinary medicine, a remarkable treatment method stands out: platelet-rich plasma (PRP) therapy. Using the body’s own healing mechanisms, this innovative approach opens new doors in regenerative medicine for our animal friends.
What is PRP?
PRP therapy involves concentrating platelets from the patient’s blood. Platelets, small blood cells that play a role in clotting and are rich in growth factors, are isolated and concentrated and used to accelerate the healing of injured tissues such as tendons and ligaments.
The Effect of PRP: Healing Our Four-Legged Friends
Veterinarians are now using PRP therapy for a variety of conditions, especially on dogs and horses. Muscle tears, ligament strains, and even more complex conditions such as major wounds or burns are showing significant improvements with PRP. It is also noted for its effectiveness in treating eye conditions such as corneal ulcers.
Today, veterinary medicine effectively uses platelet-rich plasma (PRP) therapy in many areas such as skeletal-muscular system disorders (osteoarthritis, tendon and ligament injuries, muscle damage), wound healing, post-operative healing processes, dental and oral surgery interventions, and eye diseases. This method plays an important role in the development of modern veterinary medicine, contributing to the health of animals and accelerating their healing processes.
The Science of PRP
The secret behind the success of PRP lies in the high concentration of platelets. These platelets release growth factors that help tissue repair and reduce inflammation, attracting stem cells to the site of injury. This process activates the body’s natural healing mechanisms, leading to a faster and more effective recovery.
Recent advances in PRP therapy have expanded its applications. Veterinarians can now use it not only for musculoskeletal injuries, but also in new treatment areas such as laminitis, a painful hoof disease of horses, and even traumatic brain injuries.
The effectiveness of PRP therapy depends on several factors: the concentration of platelets, the method of activation, and their correct delivery to the site of injury. Veterinary scientists are constantly improving these parameters to maximize the therapeutic benefits of PRP.
A Bright Future Ahead
As we delve deeper into the potential of PRP therapy, it is clear that this technique has great promise in veterinary medicine. By harnessing the body’s innate healing power, PRP therapy is not just a treatment, it is a revolution in the way we care for our animal companions.
PRP therapy is an example of the extraordinary advances being made in veterinary medicine. As research develops, this therapy offers a beacon of hope for pet owners and their companions, poised to transform our approach to healing and recovery in animals.
Related research articles
Alves JC, Santos A, Jorge P. Platelet-rich plasma therapy in dogs with bilateral hip osteoarthritis. BMC Vet Res. 2021 Jun 5;17(1):207. doi: 10.1186/s12917-021-02913-x.
Borş SI, Ibănescu I, Borş A, Abdoon ASS. Platelet-rich plasma in animal reproductive medicine: Prospective and applications. Reprod Domest Anim. 2022 Nov;57(11):1287-1294. doi: 10.1111/rda.14213.
McCarrel TM. Equine Platelet-Rich Plasma. Vet Clin North Am Equine Pract. 2023 Dec;39(3):429-442. doi: 10.1016/j.cveq.2023.06.007.
Meznerics FA, Fehérvári P, Dembrovszky F, Kovács KD, Kemény LV, Csupor D, Hegyi P, Bánvölgyi A. Platelet-Rich Plasma in Chronic Wound Management: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. J Clin Med. 2022 Dec 19;11(24):7532. doi: 10.3390/jcm11247532.
Sharun K, Chandran D, Manjusha KM, Mankuzhy PD, Kumar R, Pawde AM, Dhama K, El-Husseiny HM, Amarpal. Advances and prospects of platelet-rich plasma therapy in veterinary ophthalmology. Vet Res Commun. 2023 Sep;47(3):1031-1045. doi: 10.1007/s11259-022-10064-z.
Myocardial metabolism refers to the complex biochemical processes that occur within the heart muscle, or myocardium, to provide the energy needed for its continuous and vigorous contraction.
The heart has the highest metabolic demand of all our organs. Thus, myocardial metabolism is an area of interest for biochemists. The heart requires a sufficient supply of ATP to facilitate muscle contraction, sarcomere relaxation, and the active transport of ions across the cell membrane, as seen in processes like Na+/K+-ATPase. This demand for energy is contingent on the availability of oxygen. When an adequate supply of oxygen is present, glycolysis occurs aerobically, proceeding to the tricarboxylic acid (TCA) cycle and electron transport. In cases where oxygen is limited or absent, anaerobic glycolysis ensues, stopping at the pyruvate stage, which is then converted to lactate. This lactate is subsequently transported to the liver, where it is transformed into glucose through gluconeogenesis. In the cardiac muscle, the Cori cycle can become active in situations of heightened energy demand or stress, such as during exercise or in specific disease conditions.
Cori Cycle plays a vital rolein mycocardial metabolism
The Cori cycle plays a vital role in upholding energy production and glucose balance within cardiac muscle and other tissues, particularly when energy demand surges or oxygen availability is limited. In cardiac muscle, the Cori cycle comprises the subsequent stages.
Glycolysis: Glucose undergoes glycolysis, which consists of a sequence of chemical reactions within the cytoplasm of cardiac muscle cells. This process yields pyruvate and a modest quantity of ATP.
Lactate generation: In specific scenarios, such as when oxygen is scarce or during periods of elevated energy requirements, pyruvate is transformed into lactate via anaerobic glycolysis. Subsequently, lactate is released into the bloodstream.
Lactate Uptake: Lactate generated within the cardiac muscle has the capacity to be absorbed by various tissues, including the liver, skeletal muscles, or even other cardiac muscle cells. In these tissues, it can serve as an energy source or be reconverted into glucose.
Glucose Regeneration: Within the tissues that receive lactate, such as the liver, there exists a process called gluconeogenesis, which can convert lactate back into glucose. This glucose is subsequently released into the bloodstream and taken up by cardiac muscle cells, where it is utilized as an energy source, effectively concluding the Cori cycle.
The heart’s energy requirements are also influenced by the availability of substrates beyond just oxygen. About 70% of the cardiac ATP is generated through the beta-oxidation of fatty acids, which serve as the primary energy source for an adult heart. These fatty acids originate from chylomicrons and result from the hydrolysis of triglycerides by lipoprotein lipase. Carbohydrates, on the other hand, serve as the energy source for the fetal heart and for an adult heart under stressful conditions, such as during ischemia.
In unusual situations like starvation, amino acids, and ketone bodies can also be utilized to produce ATP in metabolism. Apart from substrates, there is an additional requirement for nutrients to support ATP synthesis, which includes fat-soluble vitamins such as A, D, E, and K. These vitamins are present in chylomicrons and circulating lipoproteins and are released through the action of lipoprotein lipase. Vitamin D plays a crucial role in calcium absorption from the intestines. Coenzymes like TPP (thiamine), NAD (niacin), and FAD (riboflavin), as well as electrolytes such as calcium, sodium, potassium, and chloride, are essential for ATP production as well.
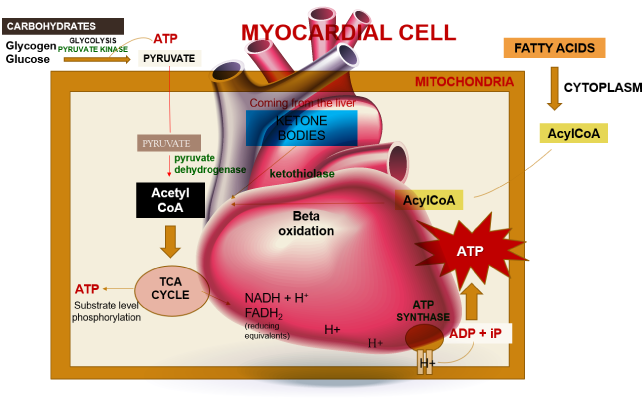
Figure 1. The three main substrates for ATP synthesis—lactate, ketone bodies, amino acids, or even acetate can be oxidized under certain circumstances.
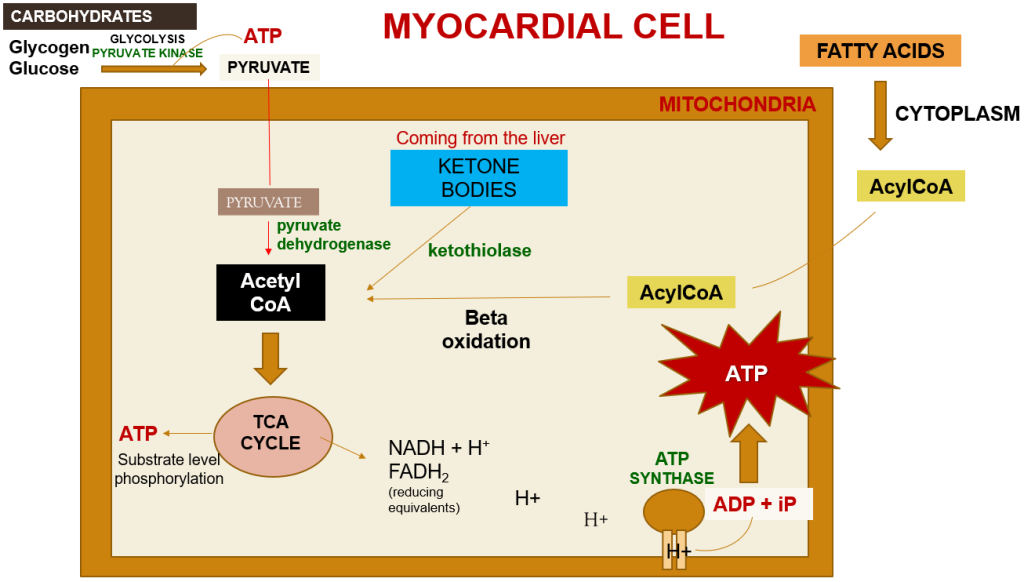
Within a myocardial cell, myocardial metabolism, glucose from the bloodstream, and glycogen stored in the myocardium go through glycolysis with the assistance of the pyruvate kinase enzyme, leading to the production of pyruvate. During this process, substrate-level phosphorylation takes place, yielding a small quantity of ATP. This pyruvate is then transported into the mitochondria, where it is converted into acetyl-CoA by the pyruvate dehydrogenase enzyme complex. Additionally, acetyl-CoA can also be produced from ketone bodies, synthesized by the liver but not utilized, and the ketothiolase enzyme plays a role in this process.
Figure 2. The nutrients needed for ATP synthesis.
Fatty acids are activated into Acyl-CoA in the cytoplasm and are then transported to the mitochondria, where they are transformed into acetyl-CoA through beta-oxidation. This acetyl-CoA enters the TCA cycle, combining with oxaloacetate. Within the TCA cycle, substrate-level phosphorylation occurs once more, generating a small amount of ATP. Besides ATP production, the TCA cycle also produces NADH+H+ and FADH2, which act as carriers of electrons. As these electrons move from one complex to another, protons enter the intramembranous space. These protons subsequently pass through ATP synthase, causing it to rotate at a high speed, which facilitates the combination of ADP and Pi to form ATP.
This newly formed ATP combines with creatine, a compound synthesized by the liver from three amino acids (glycine, arginine, and methionine). This combination results in the formation of creatine phosphate, which is synthesized in the cytoplasm of myocardial cells from creatine and ATP. Importantly, it can be rapidly converted back into ATP during periods of elevated energy demand, such as during myocardial contraction. The enzyme creatine kinase, found in the myocardium, catalyzes the transfer of a high-energy phosphate group from creatine phosphate to ADP, effectively regenerating ATP. This process offers a swift source of ATP to support myocardial contractile function during times of increased heart rate or heightened stress.
Figure 3. A summary of myocardial metabolism pathways
Several animal species are known to be susceptible to cardiac diseases.
Dogs: Certain dog breeds, including Boxers, Doberman Pinschers, Great Danes, and Cavalier King Charles Spaniels, are predisposed to specific cardiac conditions like dilated cardiomyopathy (DCM) and mitral valve disease. DCM is characterized by the weakening and enlargement of the heart, making it less efficient at pumping blood, resulting in symptoms like fatigue, breathing difficulties, and fluid retention. Mitral valve disease involves a faulty closure of the valve between the left atrium and left ventricle, leading to blood leakage, which can cause heart chamber enlargement and symptoms such as coughing, breathing problems, and heart murmurs.
Cats: Hypertrophic cardiomyopathy (HCM) is a prevalent feline cardiac ailment, particularly affecting breeds like Maine Coon, Ragdoll, and Sphynx. HCM involves thickening of the heart’s walls, reducing its pumping efficiency, and resulting in symptoms like lethargy, breathing difficulties, and irregular heartbeats.
Horses: Horses can also suffer from cardiac diseases, including atrial fibrillation, valvular heart disease, and myocarditis. Atrial fibrillation refers to an abnormal heart rhythm affecting the atria.
Birds: Certain bird species, notably parrots and pigeons, can be vulnerable to cardiovascular diseases such as heart failure and atherosclerosis.
This article has been prepared from the presentation of our student, Doğa İsmailoğlu.
References
Heinrich Taegtmeyer (2012). Chapter 15 – Cardiomyocyte Metabolism: All Is in Flux, Editor(s): Joseph A. Hill, Eric N. Olson, Muscle, Academic Press, Pages 187-202, ISBN 9780123815101 https://doi.org/10.1016/B978-0-12-381510-1.00015-6.
Kodde IF, van der Stok J, Smolenski RT, de Jong JW (2007). Metabolic and genetic regulation of cardiac energy substrate preference. Comp Biochem Physiol A Mol Integr Physiol., 146(1):26-39. https://doi.org/10.1016/j.cbpa.2006.09.014
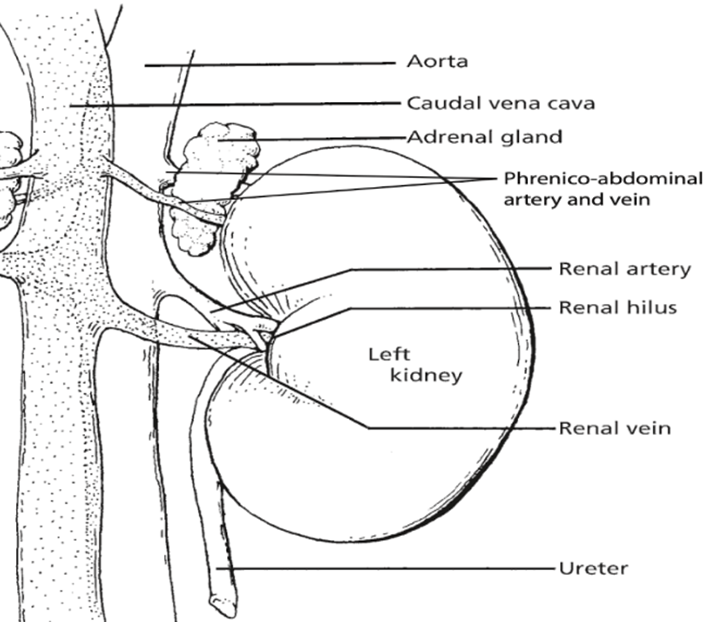
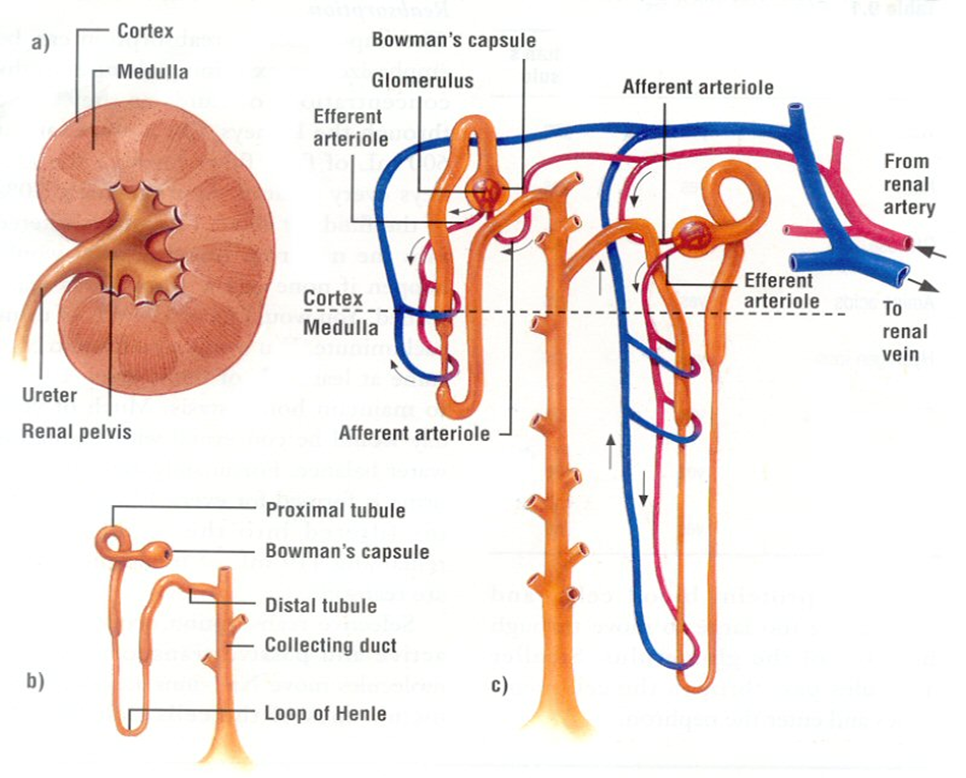
The kidneys, essential paired organs within the mammalian body, are situated symmetrically on either side of the dorsal abdomen, with the right kidney positioned on the right flank and the left kidney on the left flank.
These remarkable organs play a pivotal role in maintaining the body’s internal environment by performing the crucial function of waste product filtration from the bloodstream. This intricate process of filtration transpires as blood courses through a specialized structure known as the glomerulus. Remarkably, the specific anatomical configuration of kidneys can exhibit noteworthy variations across different species, reflecting the remarkable adaptability and diversity of life forms in the animal kingdom.
Fig. 1: Illustration of left kidney
The Nephron
The nephron, a structurally and functionally intricate unit, serves as the fundamental building block of the kidney’s physiological operations. One intriguing aspect of nephron biology is the considerable interspecies variation in their abundance. Nephrons can be broadly classified into two main categories: cortical or corticomedullary nephrons and juxtamedullary nephrons. The former, found predominantly in the outer and middle cortical regions, are characterized by the presence of glomeruli, and are associated with Henle loops that extend to the junction between the medulla and cortex, or even into the outer zone of the medulla. In contrast, juxtamedullary nephrons, as the name suggests, are situated nearer to the medulla within the renal cortex. These nephrons are distinguished by glomeruli as well and are connected to loops of Henle that extend deep into the medulla, with some Henle loops reaching as far as the renal pelvis. This structural diversity in nephron distribution and configuration underlines the remarkable adaptability of the renal system across species, allowing it to finely tune its physiological functions to meet the unique demands of each organism.
Fig. 2. Kidney’s Functional Unit Components
The kidneys, apart from their role in waste filtration, perform a multifaceted function that extends to the secretion of hormones critical for regulating blood pressure, red blood cell production, and mineral metabolism. These hormones, namely the renin-angiotensin system (RAS), erythropoietin (EPO), and 1,25-dihydroxy-vitamin D3, are pivotal for maintaining various aspects of bodily homeostasis.
The renin-angiotensin-aldosterone system (RAAS) orchestrates a complex interplay of physiological processes. It actively governs the body’s electrolyte balance, fluid equilibrium, and blood pressure regulation, thereby playing a vital role in cardiovascular health.
Erythropoietin (EPO), another significant hormone produced by the kidneys, serves as a key player in the body’s response to low oxygen levels. In situations such as high altitudes or in individuals with chronic lung disorders, EPO stimulates the bone marrow to boost the production of red blood cells. The kidneys respond to hypoxia by releasing more EPO, enhancing the oxygen-carrying capacity of the blood. In medical practice, EPO is often prescribed as a therapeutic agent to address anaemia.
The maintenance of calcium and phosphorus levels in the body is heavily influenced by 1,25-dihydroxy-vitamin D3. This hormone exerts its effects on multiple fronts, influencing the intestines to facilitate the absorption of calcium and phosphorus from dietary sources, regulating calcium deposition and resorption in bones, and controlling the production of parathyroid hormone (PTH). PTH is integral in preserving normal blood calcium levels, further emphasizing the kidneys’ role in mineral metabolism.
To execute these vital functions, kidney cells, like all cells in the body, require energy. The kidneys, in fact, stand as one of the most energy-demanding organs, surpassed only by the heart. This high energy requirement is attributed to their substantial mitochondrial content and oxygen consumption. Kidneys maintain a high resting metabolic rate, necessitating a profusion of mitochondria to meet their energy needs.
Glucose, drawn from the bloodstream, serves as a primary energy source for kidney cells. In addition to glucose, kidney cells can also utilize alternative energy substrates, including fatty acids and amino acids. This metabolic versatility is essential for the kidneys to adapt to varying energy demands and maintain their multifaceted physiological roles.
Glucose: The Key Fuel for ATP Synthesis
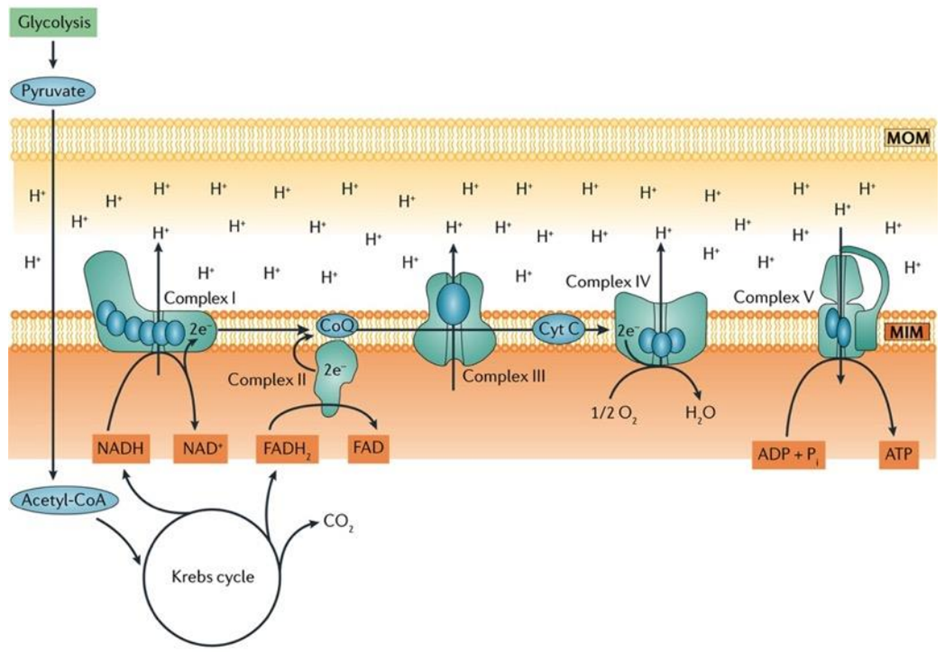
Aerobic respiration is a fundamental cellular process wherein oxygen is meticulously consumed to produce adenosine triphosphate (ATP), water, and carbon dioxide (CO2). The lion’s share of ATP generated during aerobic respiration is a direct consequence of the intricate workings of the electron transport chain (ETC) and the sequential movement of electrons within this system. The process commences with glycolysis, the initial step of aerobic respiration, which involves the conversion of glucose, the key fuel, into pyruvate.
Within the mitochondrial matrix, the pyruvate dehydrogenase complex enzymatically converts pyruvate to acetyl-CoA, a crucial substrate that fuels the tricarboxylic acid (TCA) cycle. This cycle results in the production of six molecules of nicotinamide adenine dinucleotide (NADH+H+), two molecules of flavin adenine dinucleotide (FADH2), and six molecules of carbon dioxide and water for each glucose molecule processed.
In the inner mitochondrial membrane, complex I and complex II of the ETC come into play, receiving electrons derived from NADH+H+ and FADH2, respectively. These electrons embark on an intricate journey through the electron transport chain, moving from one complex to another, until they reach complex IV. At complex IV, oxygen serves as the final electron acceptor, thus facilitating the formation of water as the terminal product.
Notably, coenzyme Q and cytochrome c, essential components situated within the inner mitochondrial membrane, play a pivotal role in mediating the transfer of electrons from complex I/II to complex III and complex III to complex IV, respectively, ensuring the seamless progression of electron flow.
Ultimately, the conversion of adenosine diphosphate (ADP) to ATP is catalyzed by ATP synthase, a dynamic enzyme complex. This enzyme harnesses the energy generated by the electron transport chain to drive the synthesis of ATP, making it a central player in the overall process of aerobic respiration, as illustrated in Fig. 3.
Fig. 3. Oxidative Phosphorylation – The Key Process for ATP Generation in Aerobic Respiration, Involving Electron Flow Through the Electron Transport Chain (ETC).
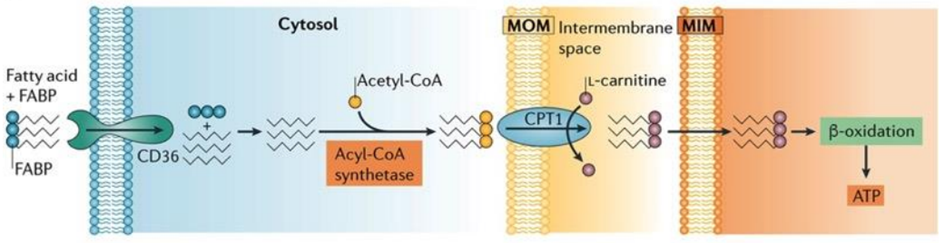
Fatty Acids: Powering the Kidney as Efficient Fuels
Proximal tubule cells rely on aerobic respiration, the most efficient mechanism for ATP production, as their energy demands are substantial due to the necessity of powering ion transport processes. Notably, fatty acids emerge as a significant energy source for proximal tubules, as a single molecule of palmitate yields a higher ATP output compared to a molecule of glucose.
Proximal tubule cells can perform one of two key functions related to fatty acids: either they synthesize fatty acids in their cytoplasm, activating them with coenzyme A before transferring them into mitochondria through the carnitine shuttle, or they can uptake fatty acids using specialized transport proteins, like platelet glycoprotein 4, also known as CD36.
In the kidney’s proximal tubule cells, the transformation of fatty acids into ATP is a complex and vital process. After the uptake or synthesis of fatty acids, these molecules need to be metabolized to yield energy. Fatty acids are first transported into the mitochondria via the carnitine shuttle, where they undergo a series of enzymatic reactions. These reactions break down the fatty acids through beta-oxidation, a process that progressively shortens the carbon chains of the fatty acids. As a result, acetyl-CoA molecules are generated from the breakdown of fatty acids. Acetyl-CoA then enters the citric acid cycle (also known as the Krebs cycle), a central metabolic pathway in mitochondria. In the citric acid cycle, acetyl-CoA is further processed to produce high-energy electrons and other intermediates. These high-energy electrons are transferred to the ETC, where they drive a series of redox reactions that ultimately lead to the production of ATP. This ATP can then be used by the kidney cells to carry out various essential functions, including the reabsorption of nutrients and the regulation of fluid and electrolyte balance within the body.
Fig. 4. Transport and activation of fatty acids within the renal proximal tubule cells. Proximal tubules have a high demand for ATP to fuel ion transport, making aerobic respiration their preferred energy generation method. Fatty acids serve as a primary energy source in these cells due to their ability to yield more ATP from a single molecule of palmitate compared to one molecule of glucose.
The Role of Amino Acids as Fuels for Kidney Cells
Amino acid transporters are abundantly present in the luminal membrane of the proximal tubule, and their reabsorption within this nephron segment is of profound importance as some of these reabsorbed amino acids can act as gluconeogenic substrates.
In the intricate landscape of the renal system, the glomerulus assumes the role of a selective filter, allowing free amino acids to pass through, while the proximal tubule primarily shoulders the responsibility of reabsorbing these amino acids. The reabsorption process itself is a multifaceted interplay involving mechanisms such as diffusion, facilitated diffusion, and sodium-dependent active transport. Furthermore, the basolateral amino acid transporters are instrumental in orchestrating the reabsorption process, each serving specific physiological functions that contribute to the overall efficiency of renal amino acid handling.
Alternatively, amino acids may also embark on an oxidative journey, subsequently entering the tricarboxylic acid (TCA) cycle at distinct junctures. A noteworthy source of energy resides in branched-chain amino acids (BCAA), comprising leucine, valine, and isoleucine.
The branch chain α-ketoacid dehydrogenase (BCKDH) complex plays a pivotal role in the metabolic pathway of BCAAs, instigating an oxidative decarboxylation reaction subsequent to the initial transamination of BCAA by branched-chain aminotransferases (BCAT), yielding branched chain α-ketoacids. Metabolites stemming from the catabolism of BCAAs traverse through the oxidative pathways of the TCA cycle, leading to the formation of succinyl-CoA or acetyl-CoA. Notably, the kidney stands as the sole tissue, apart from the heart and brown fat, where the expression and activity of both BCAT and BCKDH are distinctly observed, underscoring the kidney’s distinctive role in the robust oxidative flux of BCAAs.
Fig. 5. The metabolic linkage of amino acids to the Krebs cycle
Hormonal Regulation of Energy Metabolism in Renal Cells
Insulin: Insulin, an endocrine hormone synthesized by the pancreas, holds a pivotal role in the meticulous control of glucose metabolism. Its influence extends to the kidneys, where it stimulates the uptake and utilization of glucose, thus aiding in the maintenance of optimal blood glucose levels.
Cortisol: Cortisol, a steroid hormone produced by the adrenal glands, is instrumental in the intricate orchestration of metabolic processes and energy equilibrium. Within the renal framework, cortisol fosters gluconeogenesis—a process by which new glucose is generated—and augments the catabolism of proteins and fats, channeling these molecules toward energy production.
Epinephrine and Norepinephrine: Epinephrine and norepinephrine, both emanating from the adrenal glands and intimately associated with the “fight or flight” response, impact renal function. These hormones stimulate the release of glucose by the kidneys and augment the degradation of glycogen, the body’s stored glucose reserves, to meet the immediate energy requirements imposed during stress or demanding situations.
This article has been prepared from the presentation of our student Deniz Usta.
References
Bhargava P, Schnellmann RG (2017). Mitochondrial energetics in the kidney. Nat Rev Nephrol., 13(10):629-646. doi: 10.1038/nrneph.2017.107.
Gewin LS (2021). Sugar or Fat? Renal Tubular Metabolism Reviewed in Health and Disease. Nutrients., 13(5):1580. doi: 10.3390/nu13051580.
Reece WO, Rowe EW (2017). Functional Anatomy and Physiology of Domestic Animals. 5th edition. Wiley-Blackwell
Sahay M, Kalra S, Bandgar T (2012). Renal endocrinology: The new frontier. Indian J Endocrinol Metab., 16(2):154-5. doi: 10.4103/2230-8210.93729.
Singh S, Sharma R, Kumari M, Tiwari S (2019). Insulin receptors in the kidneys in health and disease. World J Nephrol., 8(1):11-22. doi: 10.5527/wjn.v8.i1.11.
Adipose tissue is a type of connective tissue that plays an important role in energy metabolism. It includes adipocytes. Adipocytes, which are closely associated with small blood vessels, are found singly or in groups, often within lobules surrounded by fibrous septa. Adipose tissue contains different cell types. Only one-third of the tissue consists of adipocytes. The rest form fibroblasts, macrophages, stromal cells, monocytes, and preadipocytes.
Adipose tissue is a critical regulator of systemic energy homeostasis by acting as a calorie reservoir. Under nutrient-excess conditions, adipose tissue stores excess nutrients in the form of neutral lipids, while under nutrient-deficient conditions, it provides nutrients to other tissues through lipolysis.
It is a dynamic tissue that is involved in the synthesis and storage of lipids in order to meet the energy needs of the body, and that constantly changes in volume in terms of cell number and size. Excess energy is stored in lipid droplets in the form of triglycerides.
Triglycerides, which are the most concentrated form of metabolic energy storage in humans, store twice as much energy as carbohydrates and proteins.
Simultaneously, various stromal vascular cells in adipose tissue undergo numerical and/or functional changes, contributing to the maintenance of adipose tissue’s function as an energy store and endocrine organ.
There are three types of adipose tissue
White adipose tissue (WAT) is the predominant type of fat in the human body. WAT has several biological functions, including energy storage, prevention of heat loss, protection of vital organs, and hormone secretion. Some hormones include leptin, adiponectin, and resistin.
Beige adipocyte tissue, the third and most recent type of adipocyte, can emerge in VAT in response to thermogenic stimulation, a process known as the browning of WAT. Recent research suggests that the browning of WAT deserves more attention and that therapies that target the browning of WAT can help reduce obesity. Beige adipocytes reside within WAT and expend energy to generate heat during cold exposure (called cold-induced thermogenesis). It is well known that activated beige adipose tissue can stimulate weight loss and promote resistance to obesity, making it an attractive therapeutic target tissue. Ageing is the primary risk factor for obesity and is associated with loss of beige adipose tissue, suggesting that loss of energy expenditure capacities may contribute to an obesity-prone phenotype with increasing age.
Almost every mammal has brown adipose tissue (BAT). In newborns and hibernating mammals, brown adipose tissue is especially abundant. It is also present in adults and is metabolically active, but its prevalence decreases with age. The primary function of this gland is thermoregulation. The mitochondria of BATS cells are observed to be brown in colour due to the presence of large amounts of cytochromes. It is also called the hibernating gland because these fat stores function for the animal during its awakening from hibernation.
Energy Metabolism
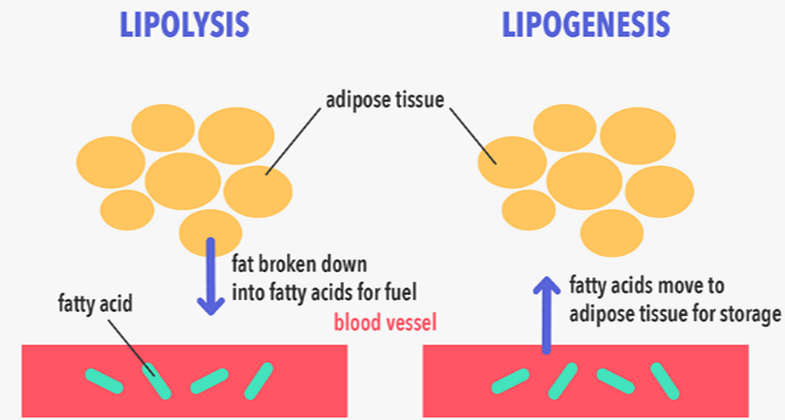
The metabolism and mobilization of lipids are under the control of adipose tissue. Lipogenesis is the process through which carbohydrates are converted into fatty acids, promoting the biosynthesis of triglycerides (TG) and the expansion of lipid droplets within adipocytes. Conversely, lipolysis breaks down TG into free fatty acids (FFA) and glycerol, which can be oxidized or released.
The uptake of circulating FFAs by the liver, muscles, and other tissues constitutes a primary pathway for lipid mobilization. Both the pathways of lipogenesis and lipolysis are highly sensitive to nutritional factors and hormones such as insulin, norepinephrine, and glucagon. As a result, the intricate regulation of these processes is essential for maintaining systemic energy homeostasis and insulin sensitivity.
Lipolysis and Lipogenesis
Lipogenesis is the term used to describe the synthesis of triglycerides and fatty acids from acetyl coenzyme A. In contrast, lipolysis involves the breakdown of triglycerides, leading to the formation of fatty acids. The key distinction between these two processes lies in their fundamental nature. Specifically, lipolysis is centered around the hydrolysis of fats and various lipid molecules, resulting in the production of fatty acids. Conversely, lipogenesis entails the creation of fatty acids and triglycerides from substrates like acetyl coenzyme A and other precursors.
Adipose tissue serves as a crucial energy storage reservoir, housing triglycerides (TGs) that are released as fatty acids through processes called lipogenesis and lipolysis, respectively.
The systemic intake of food triggers the activation of the lipogenic pathway, encouraging TG storage in adipose tissue. Conversely, fasting initiates the lipolytic pathway, prompting the breakdown of TGs and the subsequent release of fatty acids from adipose stores. This intricate balance involves lipogenesis, a process of creating fresh fatty acids from acetyl-coenzyme A (acetyl-CoA), and TG synthesis.
The metabolism of glucose generates acetyl-CoA, a pivotal component for fatty acid synthesis. This process also boosts the expression of acetyl-CoA carboxylase, the rate-controlling enzyme in lipogenesis, and triggers the release of pancreatic insulin, further propelling lipogenesis. In essence, adipose tissue functions as an energy reservoir, effectively mitigating fatty acid fluxes and averting lipotoxicity and insulin resistance. This tissue also manages the clearance of plasma TGs, averting their accumulation in other bodily tissues.
Consequently, the adipose tissue’s lipid storage capacity plays a pivotal role in systemic insulin resistance and the infiltration of lipids into organs such as the liver and muscles. On the contrary, lipolysis entails the catabolic breakdown of stored TGs within adipocytes, liberating free fatty acids and glycerol.
Starvation triggers lipolysis, yielding glycerol for hepatic gluconeogenesis and free fatty acids for oxidation, catering to the energy requirements of other organs. When fatty acids abound and carbohydrates are scarce, the liver can further metabolize fatty acids to create ketone bodies, a process termed ketogenesis, which serves as an energy source for the brain. This dynamic interplay between lipogenesis and lipolysis is pivotal for maintaining systemic energy equilibrium and insulin sensitivity. Overall, adipose tissue’s multifaceted functions underscore its significance as an energy reservoir and regulator within the body’s energy homeostasis.”
Adipose tissue acts like an endocrine organ
White adipose tissue emerges as a pivotal endocrine organ, playing a dual role in lipid storage or release and energy equilibrium by engaging in the secretion of essential adipokines. Among these, adipocytes secrete polypeptides like leptin, resistin, and adiponectin, which orchestrate a delicate balance crucial for glucose and lipid metabolism homeostasis. The intricate interplay of these adipocytokines emanating from adipocytes fundamentally contributes to sustaining optimal energy levels.
Leptin, a key player, responds to factors such as excessive energy intake, insulin levels, and glucose levels, resulting in varying production rates. Conversely, fasting, exposure to cold, β-adrenergic agonists, and testosterone lead to decreased leptin secretion. Adiponectin, a collagen-like plasma protein synthesized within adipose tissue, plays a significant role. While its concentration is higher in subcutaneous white adipose tissue, visceral white adipose tissue and hypertrophic adipocytes are inversely correlated with circulating adiponectin levels. Weight loss and periods of hunger trigger an increase in plasma adiponectin levels, which in turn activate glucose utilization within muscles. This cascade drives enhanced fatty acid oxidation in the liver and muscles, subsequently curbing glucose production due to inhibited gluconeogenesis.
The regulation of adaptive thermogenesis
Thermogenin (uncoupling protein 1, or UCP1), a distinctive molecule inherent to cold-induced thermogenesis, assumes a crucial role as it is selectively expressed within brown adipose tissue. It orchestrates a remarkable metabolic shift by diverting oxidative phosphorylation away from ATP synthesis, and channeling the energy towards heat generation instead of ATP production.
Instances of cold exposure and heightened nutritional intake trigger a surge in brown adipose tissue activity, accompanied by elevated expression levels of norepinephrine and UCP1, which emanate from the central nervous system. Notably, a repertoire of agents, including β-adrenergic antagonists, thyroid hormones, insulin, and cAMP analogues, also contribute to the augmentation of UCP1 expression.
In response to cold and nutrient availability, sympathetic nerve activity intensifies within adipose tissue. Noradrenaline binds adeptly to β-adrenergic receptors, thereby instigating a cascade of molecular signals that culminate in the hydrolysis of triglycerides. The ensuing liberation of fatty acids plays a dual role, not only energizing UCP1 but also fueling thermogenesis in cold-induced thermogenic pathways, with glucose serving as the exclusive carbon source for degradation.
Notably, the activation extends to beige cells, further enhancing the thermogenic response. Consequently, this orchestrated mechanism precipitates a surge in whole-body energy expenditure while concurrently reducing body fat mass. In essence, the interplay of these intricate processes orchestrates a metabolic symphony that elevates energy expenditure and diminishes body fat mass.
This article has been prepared from the presentation of our student Sude Sak.
References
Bódis, K, Roden, M (2018). Energy metabolism of white adipose tissue and insulin resistance in humans. Eur J Clin Invest., 48:e13017. DOI: 10.1111/eci.13017
Choe SS, Huh JY, Hwang IJ, Kim JI, Kim JB (2016). Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front Endocrinol (Lausanne), 7:30. DOI: 10.3389/fendo.2016.00030.
Mermer M, Tek NA (2017). Adipoz doku ve enerji metabolizması üzerine etkileri. Sdü Sağlık Bilimleri Enstitüsü Dergisi, 8(3): 40-46. DOI: 10.22312/sdusbed.292229
Wang Z, Wang QA, Liu Y, Jiang L (2021). Energy metabolism in brown adipose tissue. FEBS Journal, 288(12): 3647-3662. DOI: 10.1111/febs.16015.
Zhu Q, Glazier BJ, Hinkel BC, Cao J, Liu L, Liang C, Shi H (2019). Neuroendocrine Regulation of Energy Metabolism Involving Different Types of Adipose Tissues. Int J Mol Sci., 20(11): 2707. DOI: 10.3390/ijms20112707.
Metabolism is a complex process that involves various chemical reactions and pathways that occur within an organism. These reactions are essential for maintaining life and include the breakdown of food into energy, the production and storage of nutrients, and the elimination of waste products.
The regulation of metabolism is crucial for maintaining the balance between energy intake and expenditure, and hormones play a key role in this process. Hormones are signalling molecules produced by endocrine glands that regulate various physiological processes, including metabolism.
Insulin
Insulin is produced by the pancreas and is responsible for regulating glucose levels in the blood. When glucose levels rise, insulin is released to facilitate the uptake of glucose by cells for energy production or storage. Insulin also promotes the storage of excess glucose as glycogen in the liver and muscle cells.
Glucagon
Glucagon, on the other hand, is also produced by the pancreas, but it has the opposite effect of insulin. Glucagon stimulates the breakdown of glycogen in the liver to release glucose into the bloodstream when glucose levels are low. It also promotes the breakdown of fats in adipose tissue to release fatty acids for energy production.
Cortisol
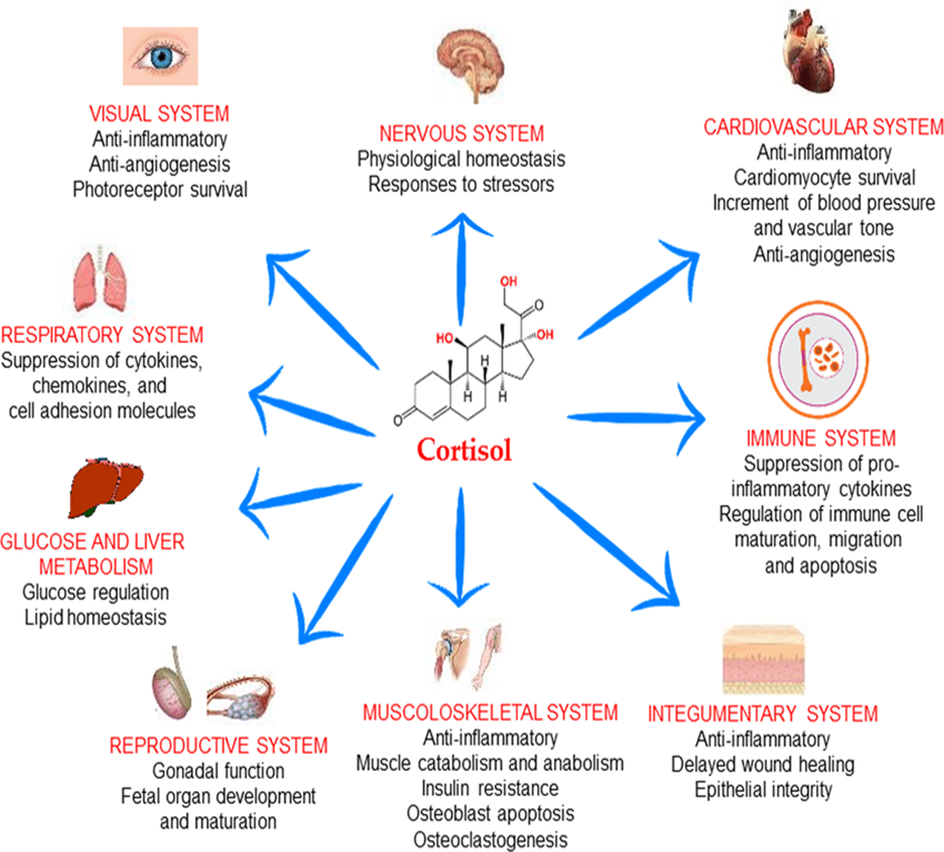
Cortisol is a steroid hormone produced by the adrenal glands in response to stress. It plays a crucial role in the metabolism of carbohydrates, proteins, and fats. Cortisol promotes the breakdown of proteins in muscle cells to release amino acids for gluconeogenesis, the production of glucose from non-carbohydrate sources. It also promotes the breakdown of fats in adipose tissue and the release of fatty acids for energy production.
Functions of Cortisol
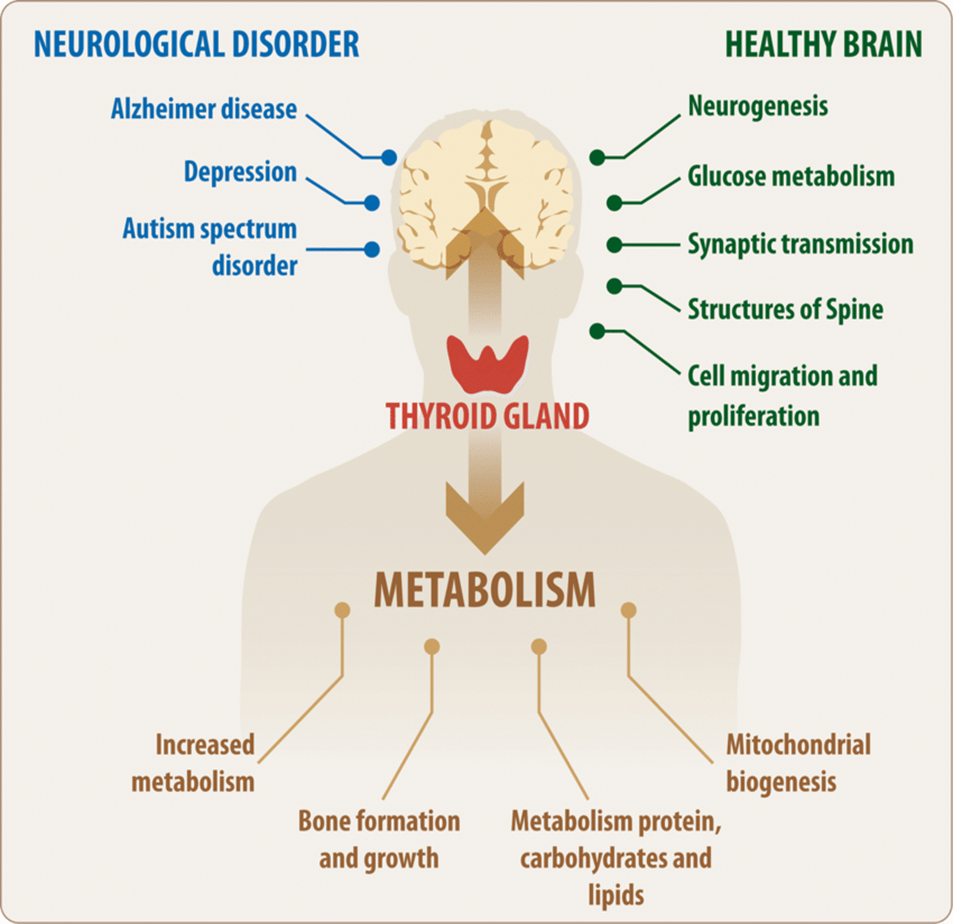
Thyroid Hormones
Thyroid hormones, produced by the thyroid gland, play a critical role in the regulation of metabolism. They increase the metabolic rate by promoting the production of ATP, the energy currency of cells. Thyroid hormones also increase the activity of enzymes involved in carbohydrate, protein, and fat metabolism
Thyroid hormones affect both physiological and pathological events.
Leptin
Leptin is another hormone that plays a crucial role in the regulation of metabolism. It is produced by adipose tissue and is involved in the regulation of energy balance. Leptin suppresses appetite and increases energy expenditure, promoting weight loss.
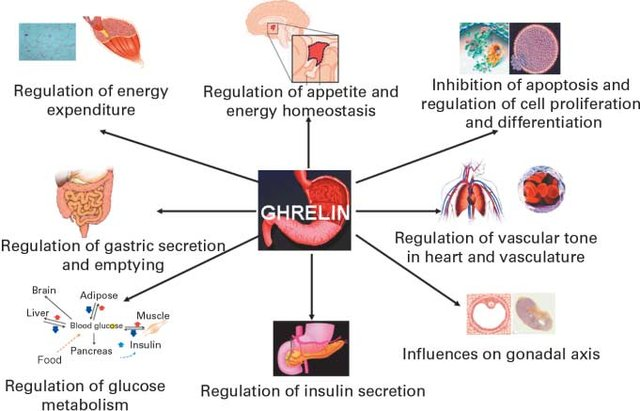
Ghrelin
Ghrelin is a hormone produced by the stomach that stimulates appetite and promotes food intake. It also plays a role in the regulation of energy balance by promoting the release of growth hormone, which increases the breakdown of fats for energy production.
Adiponectin
Adiponectin is a hormone produced by adipose tissue that regulates glucose and lipid metabolism. Adiponectin increases insulin sensitivity, promoting glucose uptake by cells for energy production. It also promotes the breakdown of fats in adipose tissue and the utilization of fatty acids for energy production.
In conclusion, hormones play a crucial role in the control of metabolism. Insulin and glucagon regulate glucose levels in the blood, while cortisol promotes the breakdown of proteins and fats for energy production.
Thyroid hormones increase the metabolic rate by promoting the production of ATP.
Leptin and ghrelin regulate appetite and energy balance, while adiponectin regulates glucose and lipid metabolism.
The proper regulation of hormones is essential for maintaining metabolic balance and overall health.
This article has been prepared from the presentation of our student Barış Ataseven.
Oxidative stress is a process that occurs due to an imbalance between reactive oxygen species (ROS) or free radicals and antioxidants in the body. ROS are molecules formed as a by-product of cellular metabolism.
Several molecules take part in the process of oxidative stress, such as superoxide anion (O2−), hydrogen peroxide (H2O2), and hydroxyl radical (•OH). As well as antioxidants like glutathione, vitamin C and E. They can cause damage to cellular components like proteins, lipids, and DNA. This can result in a variety of diseases.
Figure 1: The balance during homeostasis and the imbalance during oxidative stress.
Oxidative stress can occur in many organs, such as the liver, lungs, pancreas, intestines, and even in the eyes.
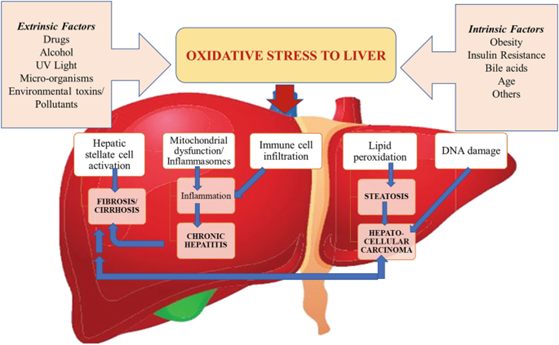
The liver plays a critical role in detoxification and metabolism, which can generate reactive oxygen species (ROS) as by-products. Excessive ROS production in the liver can result in oxidative stress and damage to liver cells. The liver produces antioxidant enzymes and molecules that can neutralize ROS. But if the liver’s antioxidant defences get overstimulated, it can lead to the accumulation of ROS which results in oxidative stress in other organs.
To counteract oxidative stress, the liver relies on antioxidant molecules such as glutathione and vitamins C and E, to neutralize ROS and prevent damage to cellular components. Some diseases related to oxidative stress in the liver are viral hepatitis, non-alcoholic fatty liver disease and liver fibrosis.
Figure 2: Factors causing oxidative stress in the liver and conditions occurring as a result of oxidative stress
The lungs are highly susceptible to oxidative stress due to their constant exposure to environmental toxins and pollutants. Oxidative stress in the lungs can lead to inflammation and damage to the lung tissue.
Pancreatic beta cells are very susceptible to oxidative stress because of their high production of reactive oxygen species and their low capacity for antioxidants. ROS can damage pancreatic beta cells. This impairs the secretion of insulin and glucose metabolism. Oxidative stress can also promote inflammation and pancreatic fibrosis, which can contribute to the development of cancer. The pancreas plays a role in the regulation of the blood sugar level by producing insulin. Dysfunction of the pancreas can lead to insulin resistance and diabetes. This can impact other organs like the kidneys, eyes and cardiovascular system.
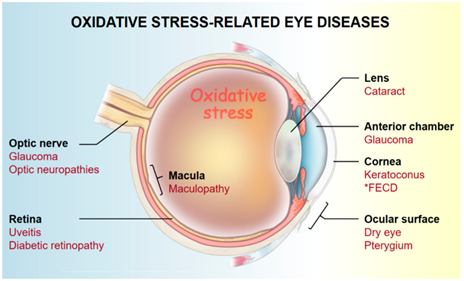
The eyes are also vulnerable to oxidative stress. This can lead to multiple diseases like cataracts, dry eyes and glaucoma. Reactive oxygen species are able to damage the lens and retina, which results in impaired vision and the increased risk of blindness. They can also damage the proteins in the lens of the eye. As an individual ages, this damage accumulates, which contributes to the development of cataract.
Figure 3: Diseases related to oxidative stress in the eyes.
Oxidative stress can also occur in the intestines. Here it can cause alteration in the microbiotics and dysfunction of the intestinal barrier. Oxidative stress can cause dysbiosis in the intestines. As a result of this, the proliferation of harmful bacteria can occur. There is a decrease in beneficial bacteria, which can further contribute to oxidative stress and inflammation in the intestines. Oxidative stress can also damage the intestinal barrier, which prevents harmful substances from entering the bloodstream. As a result of this, intestinal permeability can increase. This leads to the leakage of toxins, bacteria and other harmful substances into the bloodstream.
This article has been prepared from the presentation of our student Aline Donker.
References
Li S, Tan HY, Wang N, Zhang ZJ, Lao L, Wong CW, Feng Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int J Mol Sci. 2015 Nov 2;16(11):26087-124.
Rapa SF, Di Iorio BR, Campiglia P, Heidland A, Marzocco S. Inflammation and Oxidative Stress in Chronic Kidney Disease-Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites. Int J Mol Sci. 2019 Dec 30;21(1):263.
Betteridge DJ. What is oxidative stress? Metabolism. 2000 Feb;49(2 Suppl 1):3-8.
Hsueh, Y.-J.; Chen, Y.-N.; Tsao, Y.-T.; Cheng, C.-M.; Wu, W.-C.; Chen, H.-C. The Pathomechanism, Antioxidant Biomarkers, and Treatment of Oxidative Stress-Related Eye Diseases. Int. J. Mol. Sci. 2022, 23, 1255.
Correia AS, Cardoso A, Vale N. Oxidative Stress in Depression: The Link with the Stress Response, Neuroinflammation, Serotonin, Neurogenesis and Synaptic Plasticity. Antioxidants. 2023; 12(2):470.
Eguchi N, Vaziri ND, Dafoe DC, Ichii H. The Role of Oxidative Stress in Pancreatic β Cell Dysfunction in Diabetes. Int J Mol Sci. 2021 Feb 3;22(4):1509
Vona R, Pallotta L, Cappelletti M, Severi C, Matarrese P. The Impact of Oxidative Stress in Human Pathology: Focus on Gastrointestinal Disorders. Antioxidants (Basel). 2021 Jan 30;10(2):201.
Rogers LK, Cismowski MJ. Oxidative Stress in the Lung – The Essential Paradox. Curr Opin Toxicol. 2018 Feb;7:37-43.
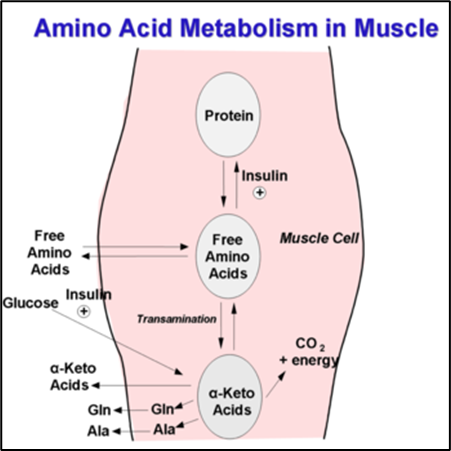
Amino acid metabolism is the collective term for the metabolic processes that produce, break down, and use amino acids. The amino groups and ammonia that are provided by the six amino acids are necessary for the synthesis of glutamine and alanine, which muscles release in enormous amounts.
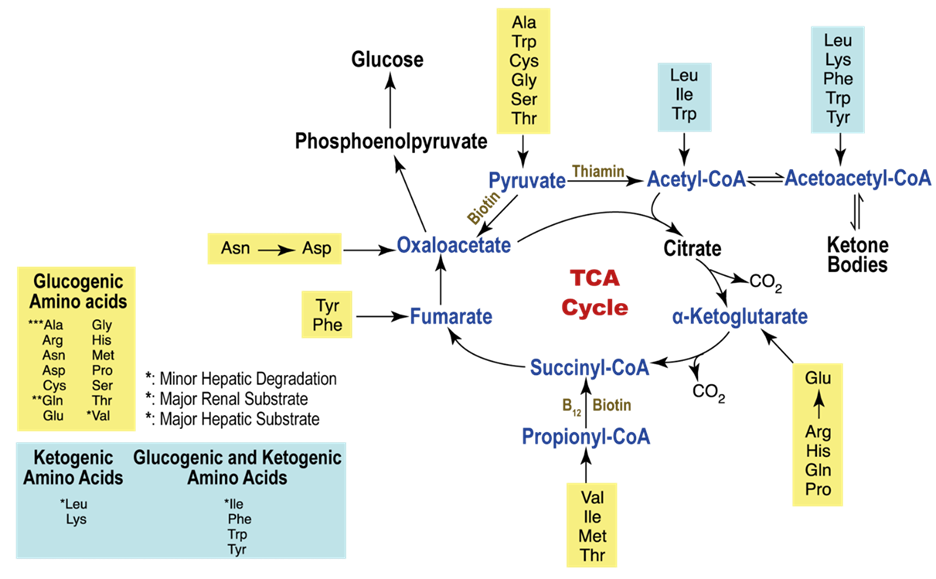
The 6 amino acids metabolized in resting muscle are Leucine, Isoleucine, Valine, Asparagine, Aspartate, and Glutamate. In addition to playing other vital roles in human metabolism, glutamine generated by muscles is an important source of energy and regulates the synthesis of DNA and RNA in the immune system and mucosal cells. Furthermore, protein synthesis and degradation are both required for the organism to maintain balance. Moreover, Acetyl-CoA can only be made by converting leucine and a portion of the isoleucine molecule, and the TCA-cycle intermediates and glutamine are synthesized using the carbon skeleton of the other amino acids.
The breakdown of amino acid carbon skeletons results in the creation of six metabolites: acetyl-CoA, acetoacetyl-CoA, pyruvate, α-ketoglutarate, fumarate, and oxaloacetate and each has a different fate in the energy metabolism. Amino acids are categorized as either ketogenic or glucogenic based on what happens to their breakdown products. Therefore Acetyl-CoA and acetoacetyl-CoA are produced by the ketogenic amino acids leucine and lysine. Many amino acids operate as direct energy-producing substrates and regulate the activity of several enzymes involved in the metabolism of glucose. Both isolated animal and human myocardium exhibit improved contractile performance as a result.
The many different ways and outcomes of Amino Acid Metabolism in Muscles.
Branched-chain amino acids (BCAAs) are the primary amino acid source for skeletal muscle anabolism and are crucial for maintaining energy balance. Skeletal muscle oxidizes the bulk of the body’s BCAAs, followed by brown adipose tissue, the liver, kidneys, the heart, and other tissues. Skeletal muscle participates disproportionately in BCAA catabolism due to the fact that BCAA transamination, the initial stage of the process, occurs predominantly there.
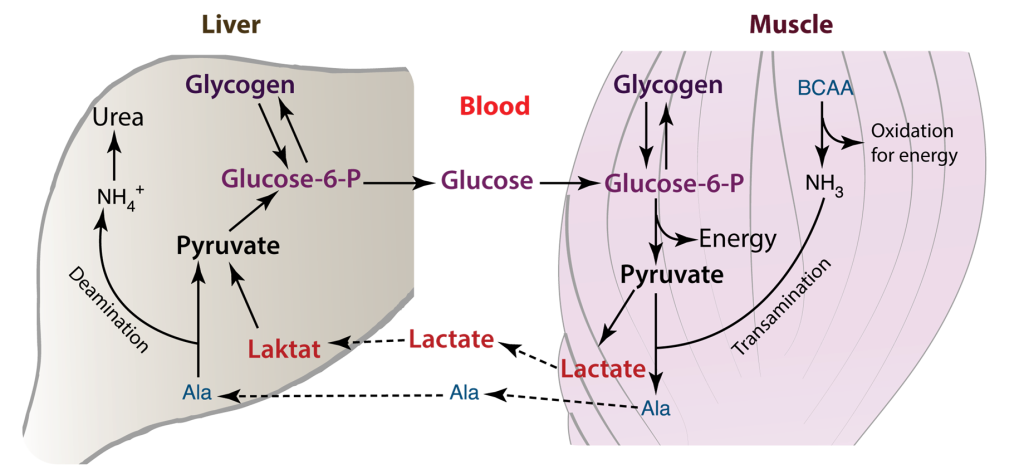
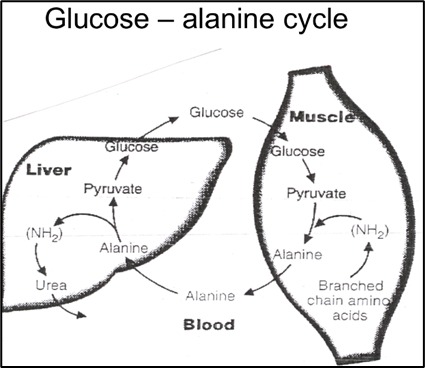
The amino acid alanine is necessary for the synthesis of proteins and it provides the central nervous system and muscles with energy. The liver uses alanine, which is secreted from skeletal muscle, as a substrate for gluconeogenesis, then the amino group of alanine is changed into urea by the urea cycle, which is then eliminated. The alanine-derived glucose produced in the liver may subsequently be able to re-enter the skeletal muscle and serve as an energy source.
Glucose – Alanine cycle and the pathways between muscle and liver.
Transamination, a chemical process in which an amino group is added to a keto acid to create new amino acids, is a crucial step in the metabolism of amino acids. The bulk of amino acids undergoes transamination during degradation. Transaminases are specific examples of enzymes that are frequently discovered as markers of potential injury to the liver cells, such as alanine aminotransferase (ALT) and aspartate aminotransferase (AST).
Aspartate transferase (AST), an essential enzyme in amino acid metabolism, is found in the liver, heart, pancreas, muscles, and other biological tissues. AST catalyzes a reaction between the amino acids aspartate and glutamate.
An enzyme called aspartate transaminase is released when your muscles or liver are injured and the main sources of AST are liver tissues, myocardium, and striated muscle. Like with all transaminases, aspartate transaminase recognizes two amino acids (Asp and Glu) with different side chains and is able to discriminate between them and bind them.
The quick oxidation of the branched-chain amino acids appears to be connected to the synthesis of alanine in muscle. Alanine is transformed into pyruvate by ALT primarily for cellular energy production moreover the enzyme ALT, which is regarded to be most closely related to the liver, is produced by the kidneys, skeletal muscle, and cardiac muscle. The intermediate metabolism of glucose and protein depends on the enzyme glutamate pyruvate transaminase, often known as alanine aminotransferase (ALT). In order to create pyruvate and glutamate, it catalyzes the reversible transamination of alanine and 2-oxoglutarate.
Leucine, alanine, and proline, three amino acids in particular, suggest they can enhance muscle repair, boost endurance, and grow muscle mass more effectively when paired with other amino acids, carbohydrates, or whey protein. Because glutamine fuels multiple cells throughout the body, it is the best recovery ingredient for all types of exercise. These acids, which are the building blocks of protein, have been demonstrated to help muscle recovery. Beyond protein, glutamine and BCAAs are two of the most important nutrients for athletes to recover and develop muscle, although BCAAs support muscle growth and prevent tiredness glutamine aids in muscle healing and rebuilding after exercise.
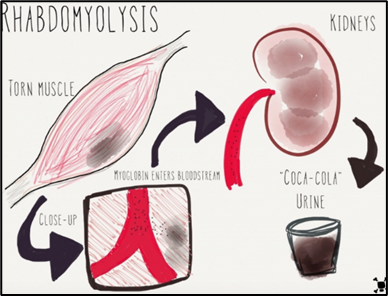
Process of Rhabdomyolysis in muscle then kidney.
For those with muscular dystrophy, defective genes hinder the body from producing the proteins required for proper muscle growth. A serious medical condition called rhabdomyolysis can be fatal or result in permanent disability. Rhabdo arises when muscle tissue is damaged because the electrolytes and proteins are discharged into the bloodstream, for instance, Myoglobin is a protein that is secreted into the bloodstream and then removed from the body by the kidneys which then produces dark-coloured urine.
This article has been prepared from the presentation of our student Tala Abdallah.
Vickie E. Baracos, Animal Models of Amino Acid Metabolism: A Focus on the Intestine, The Journal of Nutrition, Volume 134, Issue 6, June 2004, Pages 1656S–1659S
Holeček, M. “The role of skeletal muscle in the pathogenesis of altered concentrations of branched-chain amino acids (valine, leucine, and isoleucine) in liver cirrhosis, diabetes, and other diseases.” Physiological research vol. 70,3 (2021): 293-305.
Choudhary, Ankur. “General Reactions of Amino Acid Metabolism.” Pharmaguideline, https://www.pharmaguideline.com/2022/01/general-reactions-of-amino-acid-metabolism
Wagenmakers, A J. “Protein and amino acid metabolism in human muscle.” Advances in experimental medicine and biology vol. 441 (1998): 307-19.
Mann G, Mora S, Madu G, Adegoke OAJ. Branched-chain Amino Acids: Catabolism in Skeletal Muscle and Implications for Muscle and Whole-body Metabolism. Front Physiol. 2021;12:702826. Published 2021 Jul 20.
Wagenmakers, A J. “Muscle amino acid metabolism at rest and during exercise: role in human physiology and metabolism.” Exercise and sport sciences reviews vol. 26 (1998): 287-314.